Basal: Every morning, millions of people wake up, walk to the kitchen, and make coffee before they are fully conscious. They do not decide to do this. They do not weigh options. The behavior unfolds on its own, as if the brain is running a script that the person did not consciously choose to execute.
That script is stored in the basal ganglia.
These deep brain structures sit at the center of one of the most important, and least understood, aspects of human psychology: the transformation of deliberate action into automatic habit.
Understanding the basal ganglia means understanding why habits are so hard to break, why dopamine is so much more than a "pleasure chemical," and why diseases that damage these structures can rob people of the ability to move, feel motivated, or control their own behavior.
What Are the Basal Ganglia
Anatomy and Location
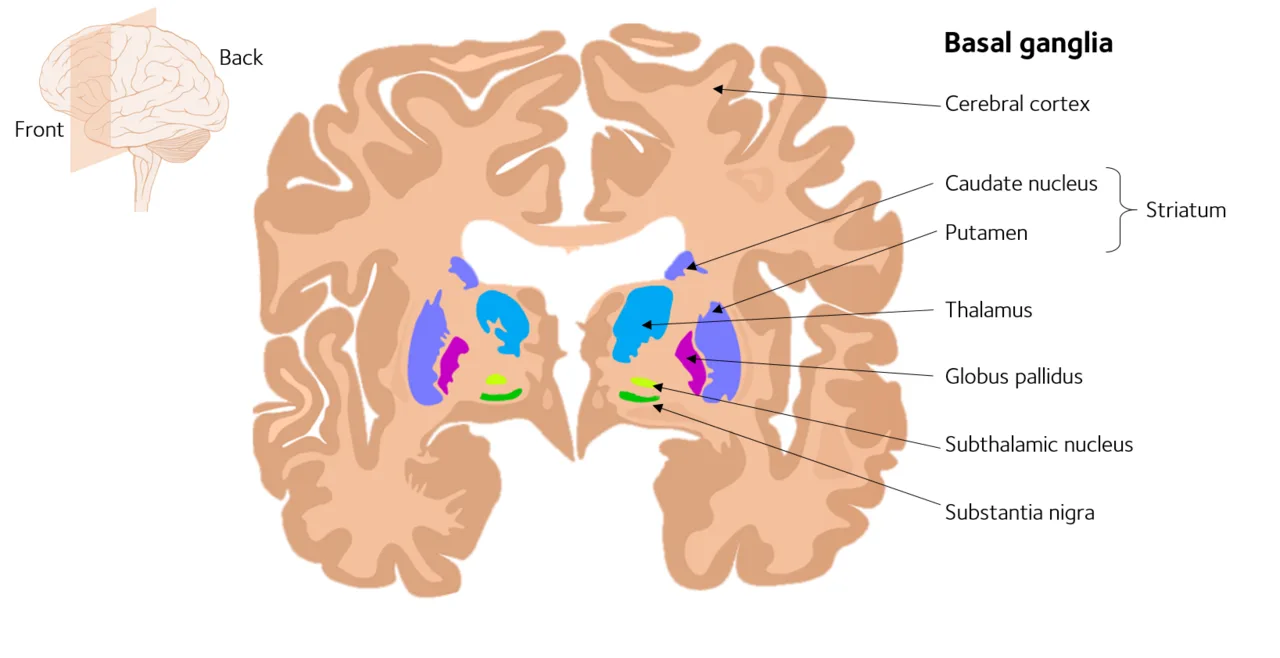
The basal ganglia are a group of interconnected subcortical nuclei, clusters of neurons below the cerebral cortex, located in the forebrain, midbrain, and upper brainstem. They are not a single structure but a functional circuit with several components:
| Structure | Location | Primary Role |
|---|---|---|
| Striatum (caudate + putamen) | Forebrain | Receives input; learns action-reward associations |
| Nucleus accumbens | Forebrain | Reward, motivation, reinforcement |
| Globus pallidus | Forebrain | Filters and modulates motor output |
| Substantia nigra | Midbrain | Produces dopamine; sends signals to striatum |
| Subthalamic nucleus | Diencephalon | Modulates movement initiation and stopping |
The striatum is the main input station of the basal ganglia, receiving signals from nearly the entire cortex as well as from dopamine-producing neurons in the substantia nigra. The globus pallidus and substantia nigra are the main output stations, projecting back to the thalamus and then to the motor cortex.
This circuit, cortex to striatum to globus pallidus to thalamus to cortex, forms a cortico-basal ganglia-thalamo-cortical loop that is central to both voluntary movement and the learning of habitual behavior.
Not Just About Movement
The basal ganglia were long described primarily as motor structures, because their most visible dysfunction, as in Parkinson's disease, produces movement problems. But decades of research have shown they are involved in much more:
- Procedural learning: mastering skills through practice
- Habit formation: automating repeated action sequences
- Reward processing: learning what predicts good and bad outcomes
- Decision-making: selecting among competing behavioral options
- Motivation: driving approach and avoidance behavior
- Cognitive control: certain aspects of working memory and attention
The basal ganglia do not just manage the body's movements, they manage which behaviors the brain selects and automates.
The Basal Ganglia and Habit Formation
How Habits Are Built
The transformation of a new behavior into a habit involves a process that neuroscientists call chunking. When you first learn a new task, say, driving a car, each component requires conscious attention: check mirrors, adjust hands, monitor speed, watch for pedestrians. The prefrontal cortex works hard.
As you repeat the task hundreds of times in consistent conditions, the basal ganglia begin to encode the entire sequence as a single unit. Activity in the prefrontal cortex decreases. Activity in the striatum increases. Eventually, the behavior runs with minimal conscious oversight.
Ann Graybiel's laboratory at MIT demonstrated this elegantly in a series of experiments with rats learning to navigate a T-maze. As rats first explored the maze, neural activity in the striatum was diffuse, firing throughout the sequence.
As training progressed and the route became habitual, striatal activity concentrated at two points: the beginning and the end of the behavioral sequence. The middle, the execution, became automatic.
"The basal ganglia, it turns out, are involved in a process that helps the brain decide when to chunk behaviors and then automates the process so that the behaviors can be enacted quickly and without deliberate thought.", Ann Graybiel, MIT
This has a critical implication: habits are not stored in consciousness. They are stored in subcortical structures that operate largely outside awareness. You do not decide to run a habit; the cue triggers it, and the behavior executes.
The Habit Loop and the Basal Ganglia
Charles Duhigg's popular framing of the "habit loop", cue, routine, reward, maps directly onto basal ganglia function:
- Cue recognition: Sensory input triggers activity in the striatum. The striatum has learned which cues predict reward.
- Routine execution: The basal ganglia release the appropriate behavioral sequence with minimal cortical involvement.
- Reward signal: Dopamine from the substantia nigra reinforces the cue-routine-reward association, strengthening the neural pathway.
Repeat the loop enough times, and the association becomes so strong that the cue alone can initiate the behavior before the person consciously registers what they are doing.
Why Habits Never Fully Disappear
One of the most counterintuitive findings from basal ganglia research is that old habits are not erased when new ones are formed. The original neural pathway remains encoded in the striatum. It becomes quieter, suppressed by competing circuits, but it is not gone.
This is why former smokers can relapse after years of abstinence when they encounter a powerful trigger. The old cue-routine-reward circuit, dormant but intact, can be reactivated by sufficiently strong cues.
The practical implication is that habit change is more accurately described as habit replacement: building a competing behavioral sequence that uses the same cue and delivers a comparable reward, eventually dominating the original circuit through repeated use.
Dopamine and Reward Prediction Error
More Than a Pleasure Chemical
Dopamine is frequently described in popular media as the brain's "pleasure chemical", the signal of enjoyment and satisfaction. This is not wrong, but it is incomplete.
The more precise description, developed through decades of neurophysiology research, is that dopamine encodes prediction error: the difference between what was expected and what actually happened.
The foundational research came from Wolfram Schultz, a neuroscientist at Cambridge. Working with macaque monkeys in the 1990s, Schultz recorded the firing patterns of dopamine neurons in the substantia nigra while the animals performed tasks with predictable rewards.
His findings were striking:
- When reward arrives unexpectedly, dopamine neurons fire strongly, a positive prediction error.
- When reward arrives as expected, dopamine neurons show no change, prediction error is zero.
- When an expected reward fails to arrive, dopamine neurons dip below baseline, a negative prediction error.
Critically, over time as the animals learned that a cue predicted the reward, the dopamine burst shifted from the reward itself to the cue. The brain stopped responding with dopamine at the moment of delivery and started responding at the moment of prediction.
Why This Matters for Behavior
This mechanism, reward prediction error, is the neural implementation of reinforcement learning, a mathematical framework developed in artificial intelligence research. The basal ganglia use prediction error signals to update their estimates of which actions lead to which outcomes, gradually sculpting behavior toward reliable reward.
The practical consequences are significant:
Variable rewards are more addictive than fixed rewards. When rewards are unpredictable, the prediction error signal is maximized, large dopamine surges occur on rewarded trials, and suppressed activity occurs on unrewarded ones.
Slot machines, social media notifications, and certain video game mechanics are engineered to exploit exactly this property.
Cues become powerful over time. Because dopamine eventually fires to the cue rather than the reward, the cue itself becomes motivationally charged. This is why certain environments, smells, or contexts can trigger cravings independent of conscious desire.
Anticipation can exceed satisfaction. Because dopamine fires to the prediction, not the outcome, wanting and liking are neurochemically distinct. You can strongly desire something and feel relatively little pleasure when you get it, a pattern common in compulsive behaviors.
Procedural Memory and Skill Acquisition
The Two Memory Systems
Neuroscientists distinguish between two broad categories of long-term memory:
Declarative memory (explicit memory) involves facts and events, things you can consciously recall and describe. "Paris is the capital of France." "I went to my sister's wedding in June." This system depends heavily on the hippocampus and medial temporal lobe.
Procedural memory (implicit memory) involves skills, habits, and conditioned responses, things you know how to do but cannot necessarily describe in words. How to ride a bicycle. How to type without looking at the keyboard. How to play a scale on the piano.
This system depends heavily on the basal ganglia (and cerebellum for timing-sensitive motor skills).
The dissociation between these systems has been demonstrated in patients with amnesia who cannot form new declarative memories but can still learn new motor skills, sometimes without any conscious awareness that learning has occurred.
The Procedural Memory Advantage
Procedural memory has a property that makes it exceptionally valuable: it is robust. Unlike declarative memory, which degrades over time without rehearsal and is susceptible to interference, procedural memory persists for years and can be retrieved even when declarative memory has failed.
People with advanced Alzheimer's disease, who may not recognize family members or remember events from earlier the same day, can still play piano pieces they learned decades ago. Veterans with severe PTSD may not be able to recall combat events explicitly but retain highly trained tactical skills automatically.
This robustness reflects the basal ganglia's architecture: once a behavioral sequence is sufficiently consolidated, it is encoded in a deeply stable form that is resistant to disruption.
The Basal Ganglia and Parkinson's Disease
What Goes Wrong
Parkinson's disease is, fundamentally, a disease of the basal ganglia. Specifically, it involves the progressive degeneration of dopaminergic neurons in the substantia nigra pars compacta, the cells that produce dopamine and send it to the striatum.
As these neurons die, the striatum receives less and less dopamine. The carefully balanced circuits that modulate motor commands become dysregulated. The result is the characteristic symptom cluster of Parkinson's:
- Resting tremor: involuntary shaking of the hands, fingers, or limbs when the body is at rest
- Rigidity: stiffness and resistance to movement in the limbs and trunk
- Bradykinesia: slowness of movement and reduced amplitude of automatic movements (arm swing, facial expression)
- Postural instability: difficulty maintaining balance, a tendency to fall
By the time symptoms appear, roughly 60-80% of the dopaminergic neurons in the substantia nigra have already been lost.
Treatment and the Dopamine Deficit
The most effective treatment for Parkinson's motor symptoms is L-DOPA (levodopa), a precursor to dopamine that can cross the blood-brain barrier (dopamine itself cannot). Once inside the brain, L-DOPA is converted to dopamine, partially restoring striatal dopamine levels and temporarily improving motor function.
Other treatments include dopamine agonists (which mimic dopamine's effects on receptors), MAO-B inhibitors (which slow the breakdown of dopamine), and deep brain stimulation (DBS), a surgical intervention that delivers electrical pulses to the subthalamic nucleus to reduce abnormal activity.
None of these treatments halt the underlying neurodegeneration. They manage symptoms while the disease progresses.
Implications for Understanding Basal Ganglia Function
Parkinson's disease has taught neuroscientists a great deal about what basal ganglia circuits actually do. Patients with Parkinson's provide a kind of natural experiment: when dopamine signaling in the basal ganglia is systematically removed, what breaks?
The answer goes beyond motor control. Parkinson's patients also show:
- Reduced ability to learn from positive outcomes (because dopamine bursts on reward are absent)
- Impaired habit acquisition (new automatic routines form slowly or not at all)
- Depression and apathy (motivational systems dependent on dopamine are disrupted)
- Cognitive slowing on tasks that require rapid action selection
This multisystem impact confirms that the basal ganglia's role in human function is far broader than movement alone.
The Basal Ganglia Across Other Conditions
Obsessive-Compulsive Disorder
OCD is associated with abnormal activity in cortico-striato-thalamo-cortical circuits, the same loops that involve the basal ganglia. Neuroimaging studies consistently find hyperactivity in the orbitofrontal cortex and caudate nucleus in OCD patients, which normalizes with successful treatment.
The compulsive behaviors of OCD can be understood as a dysregulation of the habit-formation system: behavioral sequences become locked in and cannot be suppressed by normal cortical override signals.
Addiction
Addictive drugs hijack the basal ganglia's reward prediction system. Substances like cocaine, methamphetamine, and nicotine cause massive, non-physiological dopamine releases in the striatum, particularly the nucleus accumbens.
This floods the system with artificially large prediction error signals, driving extremely strong habit formation around drug-seeking behavior.
Over time, natural rewards produce progressively less dopamine activity by comparison, a mechanism that underlies the anhedonia common in addiction.
Tourette Syndrome
Tourette syndrome involves involuntary motor and vocal tics, sudden, repetitive movements or sounds. Evidence points to abnormalities in basal ganglia-thalamocortical circuits, particularly in the inhibitory pathways that normally suppress unwanted motor outputs. When inhibition fails, unwanted behavioral sequences leak through.
Implications for Behavior Change
Working With the Habit Machine
Understanding the basal ganglia reframes behavior change from a matter of willpower to a matter of engineering.
If habits are automated sequences encoded in subcortical structures that operate largely outside conscious control, then the relevant question is not "how do I decide to behave differently?" but "how do I reshape the inputs and outputs of the habit system?"
Several evidence-based strategies follow from this framework:
Change the cue, change the habit. Because the cue triggers the entire routine automatically, altering the environment to remove or replace triggers is more effective than relying on willpower to resist them. This is why removing alcohol from the house is more effective for problem drinkers than resolving to resist it.
Keep the reward. The basal ganglia's encoding of a habit is tied to its reward. Attempting to replace a habit with a behavior that offers no comparable reward typically fails because the original cue-reward association remains stronger.
Habit substitution works best when the replacement delivers something the old behavior delivered, stimulation, stress relief, social connection.
Use repetition strategically. New habits require enough repetitions in consistent conditions for the basal ganglia to chunk them.
Research by Phillippa Lally at University College London found that on average it took 66 days for a new behavior to become automatic, but with substantial individual variation (18 to 254 days in their dataset). There is no universal "21 days" shortcut.
Implementation intentions exploit cue-routine encoding. Specifying in advance the exact cue that will trigger a desired behavior ("when I sit down with my morning coffee, I will read for 20 minutes") uses the basal ganglia's own architecture against inertia. The explicit cue-behavior pairing accelerates the encoding process.
The Limits of Conscious Override
One of the most important insights from basal ganglia research is that conscious intention is a weak override mechanism.
The prefrontal cortex can suppress habitual behavior, the research shows that inhibitory circuits from prefrontal cortex to basal ganglia do exist and do function, but they are metabolically costly, easily fatigued, and unreliable under stress.
Under conditions of high cognitive load, time pressure, hunger, or emotional arousal, conscious override of habitual behavior becomes much less effective.
This is why people tend to revert to old habits when stressed: the prefrontal cortex, already depleted, loses its grip on behavior, and the basal ganglia's default scripts run instead.
The implication is that sustainable behavior change requires making the desired behavior the automatic one, not training willpower to constantly override the default.
Research Frontiers
Optogenetics and Circuit Mapping
New tools have dramatically accelerated understanding of basal ganglia circuits. Optogenetics, a technique that uses light-sensitive proteins to activate or silence specific neurons with light, has allowed researchers to map which specific pathways within the basal ganglia control which behaviors.
Studies have used optogenetics to switch habits on and off in animal models by activating or silencing specific striatal cell populations.
Basal Ganglia and Decision-Making
Recent research has expanded the basal ganglia's recognized role in decision-making. The striatum appears to maintain a continuously updated model of the environment's reward contingencies and uses this to bias action selection in real time.
This positions the basal ganglia not just as a habit archive but as an active participant in goal-directed choices, a finding that blurs the classical distinction between "automatic" and "deliberate" behavior.
Deep Brain Stimulation Beyond Parkinson's
DBS of basal ganglia structures is being studied as a treatment for OCD, treatment-resistant depression, and Tourette syndrome.
Results are mixed but promising, and they suggest that targeted modulation of specific basal ganglia circuits may eventually offer therapeutic options for a much wider range of conditions involving dysregulated behavioral automation.
Summary
The basal ganglia are among the most consequential structures in the human brain, not because they produce conscious experience or complex reasoning, but because they quietly determine which behaviors become automatic and which rewards shape future choices.
Their role in habit formation, procedural memory, reward prediction, and motor control makes them central to understanding not just neurological disease, but the mechanisms behind learning, motivation, addiction, and the basic architecture of human behavior.
When a habit runs on autopilot, when a craving surges at a familiar cue, when a skilled pianist plays without thinking, the basal ganglia are at work. Understanding them is understanding why we do so much of what we do before we have decided to do it.
Frequently Asked Questions
What is the basal ganglia and what does it do?
The basal ganglia are a group of subcortical nuclei deep in the brain, including the striatum, globus pallidus, substantia nigra, and subthalamic nucleus, that work together to automate learned behaviors, regulate motor control, and process reward signals. They are sometimes called the brain’s habit machine because they convert deliberate, effortful actions into automatic routines over time. They also play a central role in motivation, decision-making, and the reinforcement of rewarding behaviors through dopamine signaling.
How do the basal ganglia form habits?
Habit formation in the basal ganglia occurs through a process of chunking: when a behavior is repeated consistently and reliably leads to a reward, the striatum gradually encodes the action sequence as a single automated unit. Activity in the prefrontal cortex, which manages deliberate, effortful thinking, decreases as the habit solidifies, while basal ganglia activity increases. Brain scan studies by Ann Graybiel at MIT showed that rats learning a maze showed shifting neural activity from cortex to striatum as the route became habitual.
What is reward prediction error and how does dopamine fit in?
Reward prediction error is the difference between the reward you expected and the reward you actually received. When a reward is better than expected, dopamine neurons in the basal ganglia fire strongly, reinforcing the behavior that led to it. When a reward is worse than expected, dopamine activity dips below baseline, signaling that something went wrong. This mechanism, first described by Wolfram Schultz in the 1990s, is the neural basis of reinforcement learning and explains why both wins and near-misses drive persistent behavior.
What is the connection between the basal ganglia and Parkinson's disease?
Parkinson’s disease results from the progressive loss of dopamine-producing neurons in the substantia nigra, a key component of the basal ganglia circuit. Without sufficient dopamine, the basal ganglia cannot properly modulate motor commands, leading to the characteristic symptoms of Parkinson’s: tremor at rest, muscle rigidity, slowed movement (bradykinesia), and postural instability. Treatments like L-DOPA work by replenishing dopamine levels, temporarily restoring the basal ganglia’s ability to coordinate smooth, voluntary movement.
Can you change habits by targeting the basal ganglia?
You cannot directly target the basal ganglia pharmacologically for habit change in healthy individuals without significant side effects, but behavioral strategies can work with the basal ganglia’s own mechanisms. Research suggests that habits are never truly erased, old neural pathways remain, but new routines can be built that compete with old ones. Cue manipulation (changing the environmental triggers), reward substitution, and consistent repetition of replacement behaviors all leverage the basal ganglia’s chunking process to install new automatic patterns.