Immune: In 1882, while studying starfish larvae at a makeshift laboratory in Messina, Sicily, Russian zoologist Élie Metchnikoff made an observation that would change medicine.
He inserted a rose thorn into a starfish larva and watched through his microscope as mobile cells from the larva migrated toward the foreign object and attempted to engulf it.
These were not specialized cells of a complex vertebrate immune system, they were primitive phagocytes performing what appeared to be a defensive function.
Metchnikoff had discovered phagocytosis, the process by which immune cells engulf and destroy pathogens, and in doing so had identified the ancient cellular foundation of all animal immunity.
He received the Nobel Prize in 1908 alongside Paul Ehrlich, who had taken the complementary route of studying the biochemical factors in blood serum, antibodies, that also contributed to pathogen destruction.
The debate between Metchnikoff's cellular immunity and Ehrlich's humoral immunity would preoccupy immunology for decades, before it became clear that both were essential arms of the same integrated system.
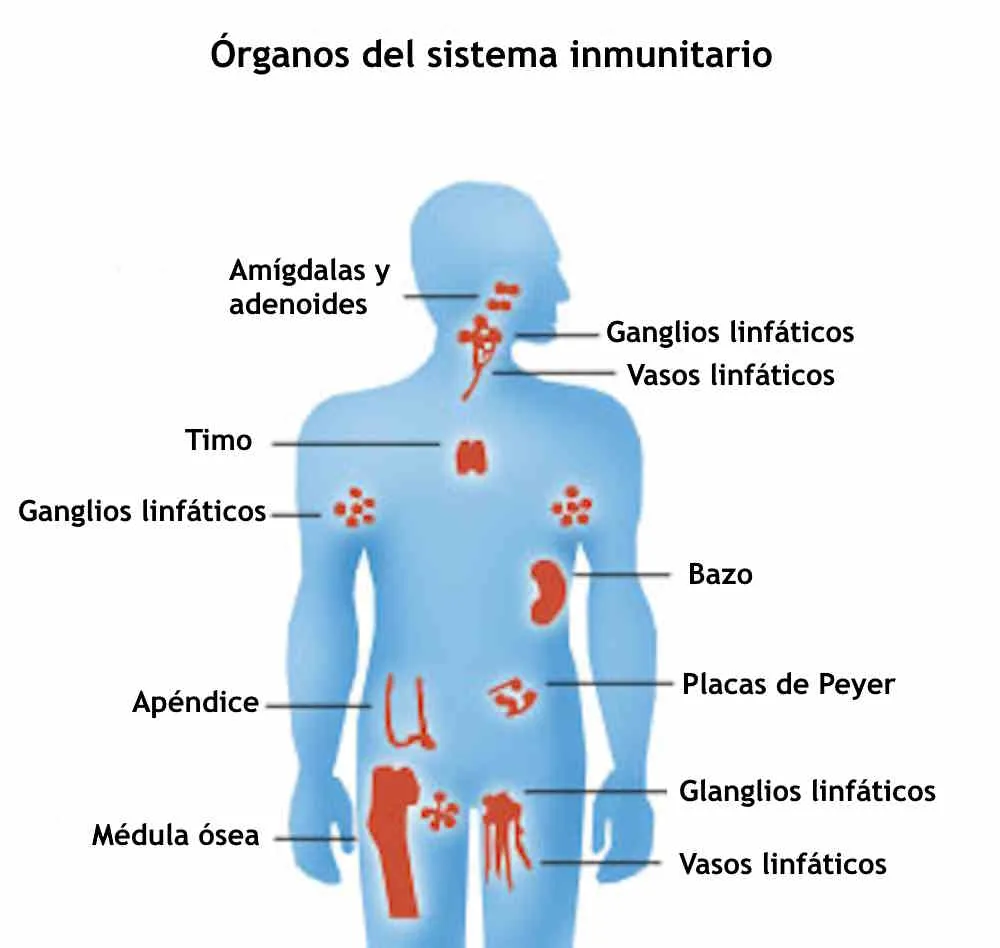
The immune system is not a single organ but an extraordinarily complex, distributed network of cells, proteins, and signaling molecules distributed throughout the body, performing the continuous work of distinguishing self from non-self, friend from foe, and damage from normal tissue turnover.
It operates on two major timescales: an innate immune system that responds within minutes to hours using pre-formed defenses against broadly conserved features of pathogens, and an adaptive immune system that develops over days to weeks by generating pathogen-specific responses that improve with each encounter and form the basis of immunological memory.
The integration of these two systems is what allows the body to mount an immediate holding action against infection while building the specific, targeted response that will ultimately clear it, and to remember that response for decades.
Understanding the immune system matters far beyond biology courses. Vaccines, cancer immunotherapy, autoimmune disease treatment, antiviral drug design, and the management of pandemic infectious diseases all depend on detailed knowledge of immunological mechanisms.
The past three decades have produced a remarkable series of Nobel Prizes in immunology, each recognizing discoveries that were not merely theoretically important but led directly to treatments saving millions of lives.
"The specific immune system is a masterpiece of evolution that can generate billions of different antibodies, each capable of recognizing a different molecular target, from a finite set of genes through a process of combinatorial diversity and somatic hypermutation.", Charles Janeway Jr., Immunobiology (2001)
Key Definitions
Innate immunity: The non-specific, rapid first-response arm of the immune system present from birth, which recognizes broadly conserved features of pathogens and mounts immediate inflammatory responses.
Adaptive immunity: The antigen-specific arm of the immune system involving B and T lymphocytes, which develops over days to weeks, generates immunological memory, and improves with repeated exposure.
Antigen: Any molecule that can be specifically recognized by an antibody or T cell receptor, typically a protein or polysaccharide on the surface of a pathogen or abnormal cell.
Antibody (immunoglobulin): A protein produced by B cells that specifically binds an antigen, neutralizing pathogens or marking them for destruction.
Phagocytosis: The process by which immune cells (phagocytes) engulf and destroy pathogens, debris, and abnormal cells.
Cytokine: A signaling protein secreted by immune cells to regulate immune responses, including interleukins (IL), tumor necrosis factor (TNF), and interferons.
MHC (major histocompatibility complex): Cell-surface proteins that present peptide fragments of antigens to T cells, enabling T cell recognition of infection or transformation.
Clonal selection: Burnet's 1957 theory that antigen exposure selects and expands the rare lymphocytes bearing receptors matching that antigen.
Immunological memory: The persistence of long-lived memory B and T cells after an immune response, enabling faster and stronger secondary responses upon re-exposure.
Checkpoint inhibitor: A drug that blocks inhibitory receptor pathways used by tumors to suppress anti-tumor T cell activity.
Key Immune Cell Types and Their Roles
| Cell type | Arm | Where produced | Primary function | Key recognition mechanism |
|---|---|---|---|---|
| Neutrophil | Innate | Bone marrow | First responder; phagocytosis of bacteria; release of toxic granules | Pattern recognition receptors (TLRs); opsonins (antibody, complement) |
| Macrophage | Innate | Bone marrow → tissues | Phagocytosis; antigen presentation to T cells; cytokine secretion; tissue resident surveillance | TLRs; scavenger receptors; Fc receptors |
| Dendritic cell | Innate / bridge to adaptive | Bone marrow → tissues | Professional antigen-presenting cell; activates naive T cells; links innate to adaptive | TLRs; C-type lectins; captures antigens in tissues, migrates to lymph nodes |
| Natural killer (NK) cell | Innate | Bone marrow | Kills virus-infected and tumor cells that downregulate MHC-I (to evade T cells) | Missing-self: absence of MHC-I signals killing; activating receptors for stress signals |
| B cell | Adaptive | Bone marrow | Produces antibodies; some become long-lived memory B cells | B cell receptor (unique antibody on cell surface); cognate antigen binding |
| CD4+ T helper cell | Adaptive | Thymus (from bone marrow precursors) | Coordinates immune response; activates B cells, CD8+ T cells, and macrophages | T cell receptor recognizing MHC-II + antigen peptide |
| CD8+ cytotoxic T cell | Adaptive | Thymus | Kills infected and tumor cells directly | T cell receptor recognizing MHC-I + antigen peptide |
| Regulatory T cell (Treg) | Adaptive | Thymus / periphery | Suppresses excessive immune activation; maintains self-tolerance; prevents autoimmunity | FoxP3 transcription factor; antigen-specific or broad suppression |
| Plasma cell | Adaptive | Lymph nodes (from B cells) | High-rate antibody secretion (up to 10,000 antibodies/second) | Derived from activated B cells; not itself antigen-driven once activated |
| Memory B and T cells | Adaptive | Lymph nodes / bone marrow | Persist for decades; enable faster, stronger secondary responses | Antigen-specific; clonally derived from primary response cells |
Physical Barriers: The First Line of Defense
Before the cellular and molecular immune system can respond to a pathogen, that pathogen must breach the body's physical barriers.
The skin is the primary external barrier, not merely a passive wall but an active microbial community, acidic pH environment, and surface rich in antimicrobial peptides (defensins) produced by keratinocytes.
The respiratory tract's mucociliary escalator, the coordinated beating of cilia lining the airways, which propels mucus and trapped particles upward toward the throat, physically removes inhaled pathogens before they can reach the lungs.
Stomach acid (pH approximately 2) is lethal to most swallowed pathogens. Mucosal surfaces throughout the gastrointestinal, respiratory, and urogenital tracts are covered by IgA antibodies, secretory immunoglobulin A, that prevent pathogen attachment.
When these physical barriers are breached, by a cut, a respiratory infection that overwhelms the mucociliary escalator, or any other route of entry, the innate immune system is the first to respond.
Innate Immunity: Pattern Recognition and Immediate Defense
Toll-Like Receptors and Pattern Recognition
The innate immune system's capacity to distinguish pathogens from self rests on pattern recognition receptors (PRRs) that detect pathogen-associated molecular patterns (PAMPs), conserved molecular features found on bacteria, fungi, viruses, and parasites but not on normal mammalian cells.
Bacterial lipopolysaccharide (LPS) on gram-negative bacterial cell walls, flagellin from bacterial flagella, double-stranded RNA produced during viral replication, and CpG DNA motifs characteristic of bacterial and viral genomes are all PAMPs recognized by PRRs.
The toll-like receptor (TLR) family is the best-characterized class of PRRs. Charles Janeway Jr.
hypothesized in 1989 that the immune system must have evolved receptors for conserved microbial patterns, proposing this as the solution to the puzzle of how innate immunity could be non-specific yet specifically distinguish self from pathogen.
Jules Hoffmann's discovery in 1996 that the Toll gene in Drosophila was essential for defense against fungal infection, and Bruce Beutler's identification in 1998 that TLR4 was the mammalian receptor for LPS, provided the experimental confirmation.
Both Hoffmann and Beutler, along with Ralph Steinman (who discovered dendritic cells), received the 2011 Nobel Prize. TLR signaling activates the master inflammatory transcription factor NF-kB, triggering production of pro-inflammatory cytokines that initiate the acute inflammatory response.
Natural Killer Cells and the Complement System
Natural killer (NK) cells patrol the circulation, identifying and killing cells that have downregulated MHC class I expression, a strategy used by some viruses and many tumors to avoid cytotoxic T cell recognition.
NK cells operate through a 'missing self' mechanism: normal cells express MHC class I, which delivers an inhibitory signal to NK cells; cells that have lost MHC class I expression receive no inhibitory signal and are killed.
This makes NK cells complementary to cytotoxic T cells, while T cells kill cells they recognize through MHC-presented antigens, NK cells kill cells that cannot be recognized.
The complement system is a cascade of plasma proteins that can be activated by pathogens through three pathways (classical, alternative, and lectin-binding), culminating in formation of the membrane attack complex, which lyses bacteria directly.
Complement fragments also function as opsonins, coating pathogens to facilitate phagocytosis, and as chemoattractants, recruiting neutrophils and other phagocytes to the site of infection.
Adaptive Immunity: B Cells and the Antibody Response
Clonal Selection and the Antibody Repertoire
Frank Macfarlane Burnet's clonal selection theory, proposed in 1957 and for which he received the Nobel Prize in 1960 (shared with Peter Medawar, who demonstrated immune tolerance), resolved a long-standing puzzle about antibody diversity.
Each B cell expresses a unique antibody-like receptor, generated during B cell development in the bone marrow through V(D)J recombination, the random joining of variable (V), diversity (D), and joining (J) gene segments encoding the antigen-binding regions of immunoglobulins.
Additional diversity is generated at the junctions between segments. The combinatorial mechanism produces an estimated pre-immune repertoire of approximately 10^11 distinct antibody specificities.
Burnet's insight was that every possible antibody specificity pre-exists in the naïve B cell pool; antigen exposure does not instruct the immune system to make a new antibody but selects among the existing repertoire, expanding the matching clones.
This selection drives clonal expansion, rapid proliferation of the matching B cell, and differentiation into either antibody-secreting plasma cells (producing up to 2,000 antibodies per second) or memory B cells that persist for decades.
After activation, B cells undergo somatic hypermutation, further diversification of their antibody variable regions through targeted mutation, followed by affinity maturation, a Darwinian selection process in germinal centers of lymph nodes that iteratively selects for B cell clones with progressively higher affinity for the antigen.
This is why secondary immune responses not only occur faster than primary responses but produce antibodies of higher quality, higher affinity and greater functional potency.
Antibody Classes and Function
The five major immunoglobulin classes reflect functional specialization for different contexts:
IgG is the dominant antibody in blood and tissues, providing long-term protection. Its four subclasses mediate complement activation, phagocytosis enhancement, and antibody-dependent cell-mediated cytotoxicity. IgG crosses the placenta, providing the fetus with maternal immunity during the first months of life.
IgM is the first antibody produced during a primary immune response and is secreted as a pentamer, making it particularly effective at complement activation and agglutination of pathogens. Its presence indicates recent or ongoing infection.
IgA protects mucosal surfaces, respiratory, gastrointestinal, and urogenital tracts, as secretory IgA (sIgA), preventing pathogen attachment. It is also the dominant antibody in breast milk, passively immunizing nursing infants.
IgE mediates allergic responses and defenses against parasitic worms (helminths). It binds mast cells and basophils; antigen crosslinking triggers histamine release, producing the symptoms of allergic reaction.
Adaptive Immunity: T Cells and Cellular Defense
Thymic Education and T Cell Diversity
T cells develop from bone marrow precursors that migrate to the thymus, where a rigorous selection process shapes the repertoire.
Like B cells, T cells generate diversity through V(D)J recombination of T cell receptor genes, the potential T cell receptor repertoire is estimated at 10^15 to 10^20 distinct specificities, substantially exceeding the B cell repertoire.
Positive selection retains T cells that can recognize self-MHC molecules; negative selection (clonal deletion) eliminates T cells that recognize self-antigens too strongly, preventing autoimmunity.
Regulatory T cells (Tregs), expressing the transcription factor FoxP3, are selected from self-reactive precursors and actively suppress other T cells to maintain peripheral tolerance.
Peter Doherty and Rolf Zinkernagel's 1974 experiments, recognized with the 1996 Nobel Prize, established MHC restriction: cytotoxic T cells infected with a virus kill only cells expressing the same MHC class I alleles, not cells of different MHC haplotypes.
This discovery established the central principle that T cells do not recognize antigens in isolation but recognize short peptide fragments of antigens presented on MHC molecules, the molecular explanation of how T cells distinguish infected from uninfected cells.
CD4+ Helper T Cells and Immunological Orchestration
CD4+ helper T cells, which recognize antigen presented by MHC class II molecules on professional antigen-presenting cells (APC), function as the orchestrators of adaptive immune responses. Distinct cytokine environments during priming direct CD4+ T cells toward different functional subsets:
Th1 cells, directed by IL-12 and IFN-gamma, produce cytokines (IFN-gamma, TNF) that activate macrophages for killing of intracellular pathogens like Mycobacterium tuberculosis and stimulate cytotoxic T cell responses.
Th2 cells, directed by IL-4, produce IL-4, IL-5, and IL-13, which promote IgE antibody production, eosinophil activation, and defense against parasitic helminths. Dysregulated Th2 responses underlie allergic diseases including asthma.
Th17 cells produce IL-17 and IL-22, defending mucosal barriers against extracellular bacteria and fungi. Inappropriate Th17 activity contributes to autoimmune diseases.
Regulatory T cells (Tregs), producing IL-10 and TGF-beta, actively suppress other immune responses to maintain self-tolerance and prevent immunopathology.
CD8+ cytotoxic T cells, which recognize antigen presented by MHC class I molecules expressed on virtually all nucleated cells, kill infected and malignant cells by releasing perforin (which forms pores in target cell membranes) and granzymes (serine proteases that enter through pores and trigger apoptosis).
HIV's strategy of specifically infecting and depleting CD4+ helper T cells is devastating precisely because it disables the orchestrating function that coordinates the entire adaptive immune response.
Vaccines and Immunological Memory
From Jenner to mRNA
Edward Jenner's 1796 demonstration that inoculation with cowpox (vaccinia) protected against smallpox was the founding experiment of vaccinology, though it was not understood mechanistically for another century.
Louis Pasteur's development of attenuated vaccines in the 1880s, weakened chicken cholera, anthrax, and rabies preparations that conferred immunity without causing disease, established the principle of using modified versions of pathogens.
Robert Koch's identification of specific causative bacteria and Pasteur's vaccines together created the germ theory framework that modern vaccinology extends.
The immunological basis of vaccine efficacy is the generation of memory B and T cells that persist after the primary response. Memory B cells express antibody receptors of higher affinity than the original naïve B cells that first responded; they also switch to IgG from the IgM-dominated primary response.
Memory T cells persist in lymph nodes, blood, and tissue sites, ready to mount secondary responses within 24-48 hours of antigen re-exposure rather than the 7-14 days required for a primary response.
The secondary response is not only faster but larger and more effective, typically clearing infection before clinical disease develops.
The mRNA vaccine platform, made clinically deployable by Katalin Karikó and Drew Weissman's 2005 discovery that pseudouridine modification of mRNA substantially reduces innate immune activation, represents a qualitative advance over previous vaccine technologies.
The Pfizer-BioNTech (BNT162b2) and Moderna (mRNA-1273) COVID-19 vaccines both encode the SARS-CoV-2 spike protein, directing cells to produce the antigen themselves.
Their approximately 95 percent efficacy against symptomatic COVID-19 in Phase 3 trials, conducted under pandemic pressure at extraordinary speed, demonstrated what the platform could deliver when regulatory, logistical, and funding barriers were lowered.
Karikó and Weissman received the 2023 Nobel Prize in Physiology or Medicine for their foundational work.
Herd Immunity and Vaccine Hesitancy
Herd immunity occurs when sufficient population immunity (through vaccination or prior infection) breaks chains of transmission, protecting individuals who cannot themselves be vaccinated.
The herd immunity threshold varies with the basic reproduction number (R0) of the pathogen: measles, with an R0 of 12-18, requires approximately 92-95 percent population immunity; COVID-19's original strain required roughly 60-70 percent.
Vaccine hesitancy, the reluctance or refusal to vaccinate despite availability, has emerged as a significant public health challenge, driven by a complex combination of safety concerns (often based on misinformation, including the debunked 1998 Wakefield paper linking MMR vaccination to autism, retracted by The Lancet in 2010 after investigation revealed data fabrication), distrust of pharmaceutical companies, political identity, and social influence.
Autoimmunity and Immunodeficiency
When the System Attacks Itself
Autoimmune diseases collectively affect approximately 5-8 percent of the Western population, with women more frequently affected than men across most conditions. Rheumatoid arthritis involves immune attack on synovial joints, with anti-CCP antibodies serving as a diagnostic biomarker.
Type 1 diabetes results from CD8+ T cell destruction of pancreatic beta cells, eliminating insulin production. Multiple sclerosis involves demyelination of central nervous system axons by autoreactive T cells and autoantibodies.
Systemic lupus erythematosus features autoantibodies against nuclear antigens, causing multi-organ damage through immune complex deposition.
The hygiene hypothesis, proposed by David Strachan in a 1989 BMJ paper reporting an inverse relationship between older siblings and hay fever, suggested that reduced childhood infections may impair tolerance mechanisms.
Graham Rook's refinement, the 'old friends' hypothesis, identifies the relevant partners as not acute pathogens but the microbial commensals, parasites, and environmental organisms present throughout human evolutionary history that co-regulate immune responses.
The dramatic rise in allergic and autoimmune diseases in industrialized countries over the past century coincides with reduced microbial diversity exposure, antibiotic use, and cesarean section deliveries that alter the infant microbiome.
Cancer Immunology: Surveillance, Evasion, and Immunotherapy
Checkpoint Inhibitors and the 2018 Nobel Prize
The concept of immune surveillance, that the immune system continuously monitors for and eliminates nascent tumors, was proposed by Burnet and Lewis Thomas in the 1950s.
Clinical evidence suggests it is real: immunosuppressed transplant recipients have substantially elevated cancer rates, and tumor-infiltrating lymphocytes correlate with better prognoses in many cancers.
Yet tumors clearly evade surveillance, using multiple mechanisms: downregulation of MHC class I to hide from cytotoxic T cells; production of immunosuppressive cytokines (TGF-beta, IL-10); exploitation of inhibitory checkpoint pathways.
James Allison identified CTLA-4 as a negative regulatory receptor on T cells, a brake on T cell activation, and demonstrated in 1996 that antibody blockade of CTLA-4 enhanced anti-tumor immune responses in mice.
Tasuku Honjo identified PD-1 as a T cell inhibitory receptor in 1992 and showed that PD-1 knockout mice developed autoimmune disease, confirming its immune-braking function.
Subsequent work demonstrated that many tumors express PD-L1, the ligand for PD-1, as an 'off switch' to paralyze infiltrating T cells.
Allison and Honjo received the 2018 Nobel Prize. Ipilimumab (anti-CTLA-4), pembrolizumab and nivolumab (anti-PD-1) have produced durable remissions in melanoma, non-small cell lung cancer, renal cell carcinoma, and many other tumor types.
The 2023 Phase 2b trial of mRNA-4157 (individualized cancer vaccine encoding neoantigens specific to each patient's tumor mutations) combined with pembrolizumab in resected high-risk melanoma showed a 49 percent reduction in recurrence or death versus pembrolizumab alone, the first significant clinical evidence that personalized mRNA cancer vaccines can substantially improve outcomes when combined with checkpoint inhibition.
See also: How Vaccines Work, How the Immune System Learns, How Cancer Develops
Sources & Further Reading
- Janeway, C. A., Jr., Travers, P., Walport, M., & Shlomchik, M. J. (2001). Immunobiology: The Immune System in Health and Disease (5th ed.). Garland Science.
- Metchnikoff, E. (1892). Lecons sur la pathologie comparee de l'inflammation. Masson.
- Burnet, F. M. (1957). A modification of Jerne's theory of antibody production using the concept of clonal selection. Australian Journal of Science, 20, 67–69.
- Janeway, C. A., Jr. (1989). Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harbor Symposia on Quantitative Biology, 54(1), 1–13.
- Hoffmann, J. A., Kafatos, F. C., Janeway, C. A., & Ezekowitz, R. A. (1999). Phylogenetic perspectives in innate immunity. Science, 284(5418), 1313–1318.
- Doherty, P. C., & Zinkernagel, R. M. (1974). Restriction of in vitro T cell-mediated cytotoxicity in lymphocytic choriomeningitis within a syngeneic or semiallogeneic system. Nature, 248(5450), 701–702.
- Karikó, K., Buckstein, M., Ni, H., & Weissman, D. (2005). Suppression of RNA recognition by Toll-like receptors: The impact of nucleoside modification and the evolutionary origin of RNA. Immunity, 23(2), 165–175.
- Allison, J. P. (1996). Tumor immunity and cancer therapy. Philosophical Transactions of the Royal Society B, 353, 1271–1278.
- Strachan, D. P. (1989). Hay fever, hygiene, and household size. BMJ, 299(6710), 1259–1260.
- Wolchok, J. D., et al. (2022). Long-term outcomes with nivolumab plus ipilimumab in advanced melanoma. New England Journal of Medicine, 386(2), 151–160.
- Moderna and BioNTech. (2020). Phase 3 trial results for mRNA COVID-19 vaccines. New England Journal of Medicine, 383, 2603–2615 and 384, 403–416.
- Weber, J. S., et al. (2024). Individualized neoantigen therapy mRNA-4157 (V940) plus pembrolizumab after resection of high-risk melanoma (KEYNOTE-942). Lancet, 403(10427), 632–644.
Frequently Asked Questions
What is the difference between innate and adaptive immunity?
Innate immunity is the immune system’s first-response layer, fast, non-specific, and present from birth. When a pathogen breaches the body’s physical barriers, innate immune cells like neutrophils, macrophages, and natural killer cells respond within minutes to hours using pattern recognition receptors (PRRs) that detect conserved features of pathogens called pathogen-associated molecular patterns (PAMPs). The innate response does not require prior exposure to a specific pathogen and cannot learn or improve with repeated encounters. Its primary roles are to immediately contain infection, initiate inflammation, and activate the adaptive immune system. Jules Hoffmann and Bruce Beutler received the 2011 Nobel Prize in Physiology or Medicine for their discovery of toll-like receptors, a key class of PRRs, in flies and mice respectively, work that had been theoretically anticipated by Charles Janeway Jr.’s 1989 hypothesis about ‘pattern recognition.’ Adaptive immunity develops over days to weeks after infection and involves the activation of B and T lymphocytes, each bearing unique receptors capable of recognizing a specific antigen. The adaptive response is both highly specific to the invading pathogen and capable of forming immunological memory, the basis of vaccination. The two systems are deeply interconnected: innate immune signaling initiates and shapes adaptive responses, while adaptive immune products (antibodies, cytokines) enhance innate effector functions. This integration means that defects in either arm produce broad vulnerability to infection.
How do B cells and antibodies work?
B lymphocytes are the antibody-producing cells of the adaptive immune system. Each B cell expresses a unique B cell receptor (BCR), a surface immunoglobulin generated through a process of V(D)J recombination, in which gene segments encoding variable, diversity, and joining regions are randomly joined during B cell development in the bone marrow. This combinatorial mechanism, together with further diversification through the joining process itself, generates an estimated repertoire of approximately 10^11 distinct antibody specificities, enough to recognize virtually any molecular structure. When a B cell encounters an antigen that binds its BCR, and receives appropriate co-stimulation from helper T cells, it becomes activated and undergoes clonal expansion, rapid multiplication of copies of itself. Frank Macfarlane Burnet’s clonal selection theory (1957), for which he received the Nobel Prize in 1960, correctly predicted this mechanism: before exposure, every possible antibody exists at low levels in the naïve repertoire; antigen exposure ‘selects’ the matching clone and drives its expansion. Activated B cells differentiate into plasma cells that secrete antibodies at high rates (up to 2,000 antibody molecules per second), or into memory B cells that persist for decades and can be rapidly recalled by subsequent exposure. Antibodies work through several mechanisms: neutralization (blocking pathogen binding sites), opsonization (coating pathogens to facilitate phagocytosis), and complement activation (triggering an enzyme cascade that lyses pathogens directly). The five antibody classes (IgG, IgM, IgA, IgE, IgD) have distinct structures and functions: IgG is the main blood antibody providing lasting protection; IgM is the first antibody produced in a response; IgA protects mucosal surfaces; IgE mediates allergic reactions.
What are T cells and how do they recognize pathogens?
T lymphocytes mature in the thymus, where they undergo selection processes that eliminate cells recognizing self-antigens too strongly (which could cause autoimmunity) and positively select cells that can recognize antigens in the context of the body’s own major histocompatibility complex (MHC) molecules. Peter Doherty and Rolf Zinkernagel received the 1996 Nobel Prize for the discovery of MHC restriction, the finding that cytotoxic T cells recognize viral peptides only when presented by the appropriate MHC class I molecules, establishing the fundamental principle of antigen presentation. T cells fall into two broad functional categories. CD4+ helper T cells recognize antigen presented by MHC class II molecules, which are expressed on professional antigen-presenting cells including dendritic cells, macrophages, and B cells. Helper T cells provide essential co-stimulatory signals for B cell activation, direct the type of immune response through cytokine secretion, and orchestrate the overall adaptive response. The CD4+ helper T cell population itself divides into functional subsets including Th1 cells (promoting cell-mediated immunity against intracellular pathogens), Th2 cells (promoting humoral responses and defense against parasites), Th17 cells (defending mucosal barriers), and regulatory T cells (Tregs, which suppress immune responses to prevent autoimmunity). CD8+ cytotoxic T cells recognize antigen presented by MHC class I molecules, expressed on virtually all nucleated cells, and kill infected or transformed cells by releasing perforin and granzymes. Their importance in cancer surveillance is substantial: tumors that downregulate MHC class I expression can escape cytotoxic T cell killing, a mechanism that checkpoint inhibitor therapies are designed to counter.
How do vaccines work and why was the mRNA vaccine a breakthrough?
Vaccines exploit the adaptive immune system’s capacity for immunological memory. Edward Jenner’s 1796 observation that milkmaids who had contracted cowpox appeared protected against smallpox was the first systematic demonstration of this principle. Louis Pasteur developed the concept of attenuated vaccines in the 1880s, using weakened or killed pathogens to stimulate immunity without causing disease, and coined the term ‘vaccination’ in honor of Jenner (from ‘vacca,’ Latin for cow). Modern vaccines present the immune system with antigens, fragments of pathogens or whole attenuated organisms, that activate B and T cell responses. After the primary response, long-lived memory B and T cells persist, sometimes for decades or for life. Upon subsequent exposure to the actual pathogen, these memory cells enable a dramatically faster and stronger secondary response that clears infection before it can cause serious disease. The mRNA vaccine technology, developed through decades of research by Katalin Karikó and Drew Weissman, for which they received the 2023 Nobel Prize in Physiology or Medicine, solved a fundamental problem: unmodified mRNA injected into cells triggers a strong innate immune response that destroys it before it can produce sufficient protein. Karikó and Weissman’s 2005 paper demonstrated that modifying specific nucleosides within the mRNA, replacing uridine with pseudouridine, dramatically reduced this innate response while preserving the mRNA’s protein-coding function. The Pfizer-BioNTech and Moderna COVID-19 vaccines, both using this modification, achieved approximately 95 percent efficacy against symptomatic COVID-19 in Phase 3 trials, a result that would not have been possible without this fundamental discovery. The mRNA platform’s advantage is speed, the sequence can be updated without rebuilding the entire manufacturing process, and its potential extends to cancer vaccines, which are now in clinical trials.
What causes autoimmune diseases and how does the immune system avoid attacking itself?
Autoimmune diseases occur when the immune system mounts a response against the body’s own tissues. Multiple mechanisms of self-tolerance normally prevent this. Central tolerance occurs during lymphocyte development in the thymus and bone marrow: T and B cells that react too strongly to self-antigens presented during development are deleted by a process called clonal deletion. The transcription factor AIRE (autoimmune regulator), expressed in thymic epithelial cells, enables presentation of a broad range of self-antigens during T cell development, expanding the scope of central tolerance. Peripheral tolerance mechanisms, including regulatory T cells and the requirement for co-stimulatory signals for full lymphocyte activation, provide additional checkpoints. Autoimmunity results when these mechanisms fail. In rheumatoid arthritis, the immune system attacks synovial joints, detectable through anti-CCP (anti-cyclic citrullinated peptide) antibodies. In type 1 diabetes, CD8+ cytotoxic T cells destroy the insulin-producing beta cells of the pancreas. In multiple sclerosis, T cells attack myelin sheaths around neurons. In systemic lupus erythematosus, autoantibodies against DNA and nuclear proteins cause multi-organ damage. David Strachan’s 1989 paper in the BMJ, reporting that children with more older siblings had lower rates of hay fever, proposed what became known as the hygiene hypothesis: reduced childhood exposure to infections may inadequately train regulatory pathways. Graham Rook’s ‘old friends’ hypothesis refined this, proposing that the relevant exposures are not acute pathogens but the microbial commensals and environmental organisms with which humans co-evolved and which help calibrate immune regulation. HIV/AIDS illustrates the consequences of adaptive immune failure from the opposite direction: the virus preferentially infects CD4+ helper T cells, and AIDS is clinically defined as CD4+ cell counts below 200 cells per microliter, at which point the immune system can no longer defend against opportunistic infections.
What is cancer immunotherapy and why did checkpoint inhibitors win a Nobel Prize?
The immune system surveys the body for abnormal cells, including tumors, through a process theorized by Frank Macfarlane Burnet and Lewis Thomas in the 1950s as ‘immune surveillance.’ Tumors, however, evolve mechanisms to evade immunity, including downregulation of MHC class I molecules to avoid cytotoxic T cell recognition, secretion of immunosuppressive cytokines, and exploitation of checkpoint pathways that normally prevent autoimmune overactivation. Checkpoint inhibitor therapy reverses this evasion by blocking the molecular ‘brakes’ that tumors exploit to suppress anti-tumor immune responses. James Allison identified CTLA-4 as a negative regulatory receptor on T cells in the 1990s and demonstrated that blocking it with antibodies enhanced anti-tumor immunity in mice. Tasuku Honjo identified PD-1 as another checkpoint receptor, and subsequent research showed that the PD-1/PD-L1 pathway was heavily exploited by tumors to suppress cytotoxic T cell activity. Allison and Honjo received the 2018 Nobel Prize for these discoveries, which led directly to drugs (ipilimumab targeting CTLA-4; pembrolizumab and nivolumab targeting PD-1) that have produced durable remissions in patients with melanoma, lung cancer, renal cell carcinoma, and many other tumor types that were previously rapidly fatal. CAR-T cell therapy represents a different approach: patient T cells are extracted, genetically engineered to express chimeric antigen receptors targeting tumor-specific surface proteins, and reinfused. In hematological malignancies including certain leukemias and lymphomas, CAR-T therapy has achieved complete remissions in patients with no other options. Personalized mRNA cancer vaccines, encoding neoantigens specific to an individual patient’s tumor mutations, showed a 49 percent reduction in recurrence or death in a Phase 2b trial of mRNA-4157 combined with pembrolizumab for resected high-risk melanoma, published in 2023, suggesting that the mRNA platform may produce a second revolution in oncology following its role in COVID-19.
What is the inflammatory response and when does it become harmful?
Inflammation is the immune system’s coordinated local response to tissue injury or infection, evolved to contain damage, eliminate pathogens, and initiate repair. The cardinal signs, redness, heat, swelling, and pain, were described by the Roman physician Aulus Cornelius Celsus in the 1st century CE and remain clinically relevant. The molecular underpinnings involve cytokines, signaling proteins secreted by immune cells, including tumor necrosis factor alpha (TNF-alpha), interleukin-1 (IL-1), and interleukin-6 (IL-6), which coordinate recruitment of neutrophils and monocytes to the site of infection, increase vascular permeability, and trigger systemic responses including fever. Fever, counterintuitively, appears to be an adaptive response: elevated temperature impairs pathogen replication while enhancing multiple aspects of immune function. The inflammasome, a multiprotein complex that activates IL-1 processing, plays a central role in sensing cellular damage and mounting the sterile inflammatory response that underlies many chronic diseases. Chronic inflammation occurs when this acute response fails to resolve, producing sustained tissue damage in conditions including atherosclerosis, type 2 diabetes, inflammatory bowel disease, and neurodegeneration. COVID-19 produced a particularly instructive case of harmful hyperinflammation: in a subset of patients, SARS-CoV-2 infection triggered a ‘cytokine storm’, dysregulated overproduction of pro-inflammatory cytokines, that caused the immune system to destroy lung tissue faster than the virus itself. This mechanism, rather than direct viral pathology, was responsible for many of the most severe COVID-19 outcomes and led to the use of dexamethasone and IL-6 receptor antagonists as life-saving treatments. Long COVID, affecting an estimated 10-20 percent of people after acute infection, involves immune dysregulation that may include viral persistence, autoantibody production, and sustained activation of innate immune pathways.